
Orchids and milkweeds export pollen not as independent grains but as coherent packages — pollinia bound into pollinaria. This convergent device makes the individual fruit a plausible, and often observed, unit of low effective sire number.
A tempting inference is that packaged transfer therefore guarantees single paternity, and that single paternity erodes genetic diversity. This work argues the opposite: single-sire capsules sit inside polygamous, well-mixed plants, and the systems usually invoked to support a diversity-loss story in fact refute it. We replace loaded shorthand with three precise quantities: capsule-level single paternity, effective sire number per capsule, and male reproductive-success variance.
The biological problem
A pollinarium is one of the most elaborate pollen-dispersal devices in flowering plants. In Orchidaceae and in the milkweeds (Asclepiadoideae, within Apocynaceae), pollen is aggregated into pollinia and coupled mechanically to the pollinator, so that an entire male gamete payload moves as a single positionally controlled unit.
The two systems are a textbook case of convergence rather than homology: orchid pollinaria use a rostellum / viscidium / stipe-or-caudicle system, whereas asclepiad pollinaria use corpuscula, translator arms, guide rails and stigmatic chambers. The shared functional outcome — accurate, package-based transport — is built from different tissues and different developmental programmes.
Pollinaria as the zero-carryover endpoint
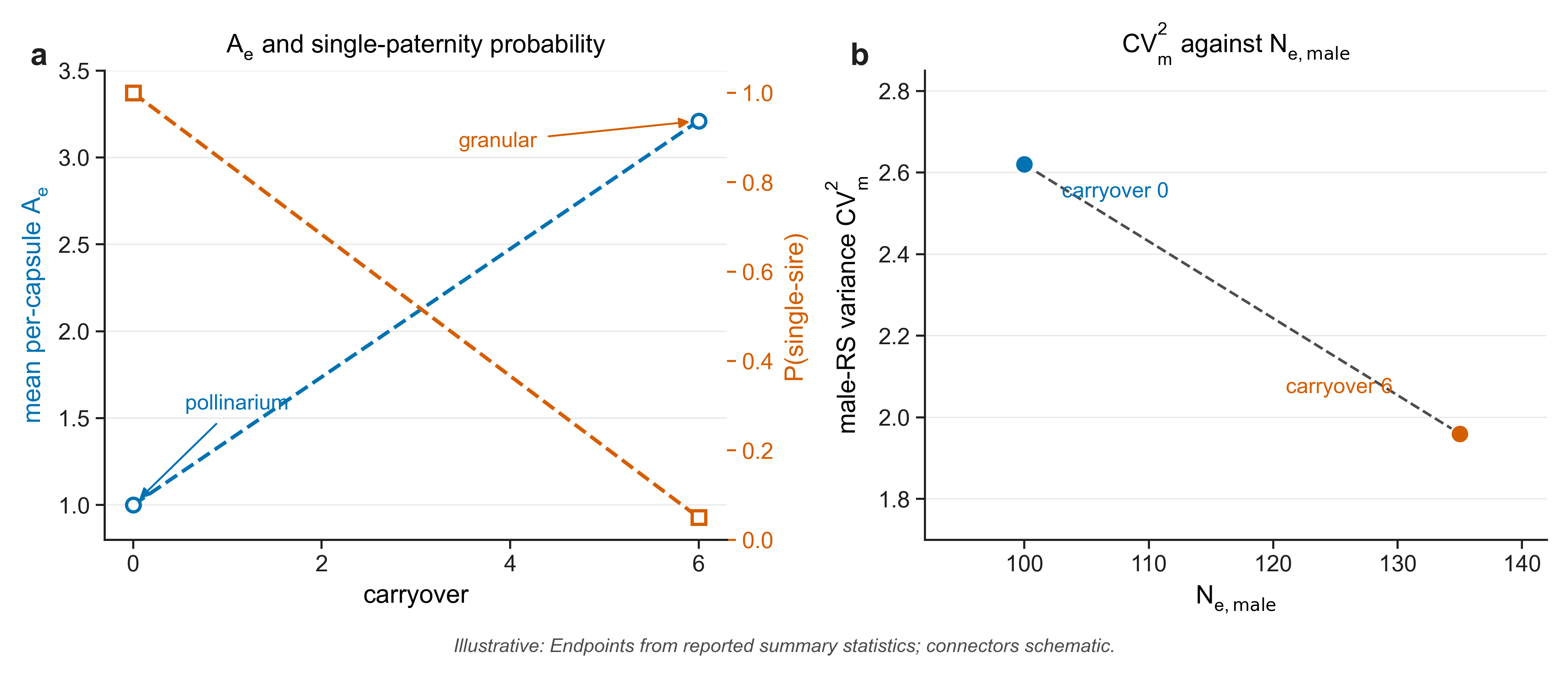
Rather than treating packaged transfer as a phenomenon of a different kind, we place it on the existing pollen-carryover / sire-profile continuum. As carryover increases from zero, the probability of a single-sired capsule falls smoothly from near 1.00 toward 0.05 and effective sire number climbs. The pollinarium simply occupies the limit of that axis.
Key reframing. Packaged transfer is quantitatively continuous with loose-pollen systems, not categorically separate. Its distinctiveness is that it pushes one knob — pollen carryover — to its limit, making the capsule the natural unit of single paternity.
The model
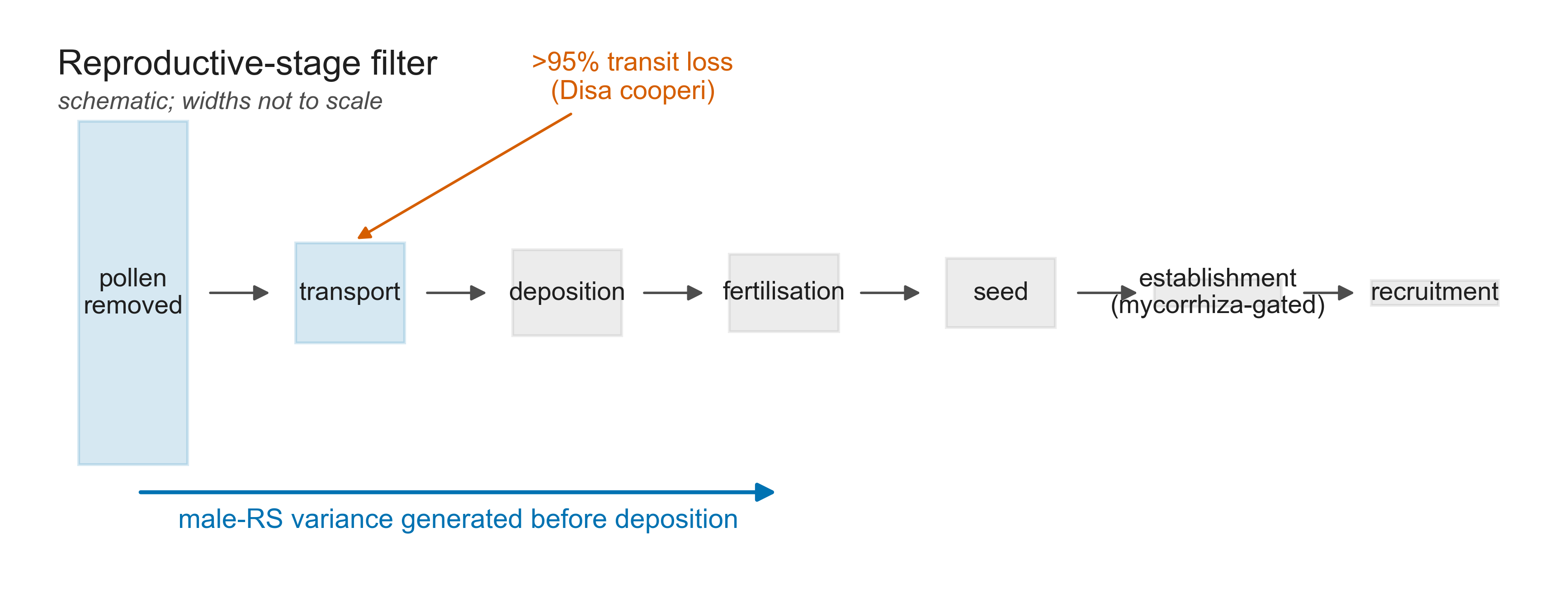
We formalise the per-capsule expectation as an emergent property of a deposition process and test it with an individual-based, forward-time simulation run on identical demography. Every pollinarium regime is compared against a matched granular multi-sire null, so that no diversity effect can be attributed to packaging when it is really an effect of demography.
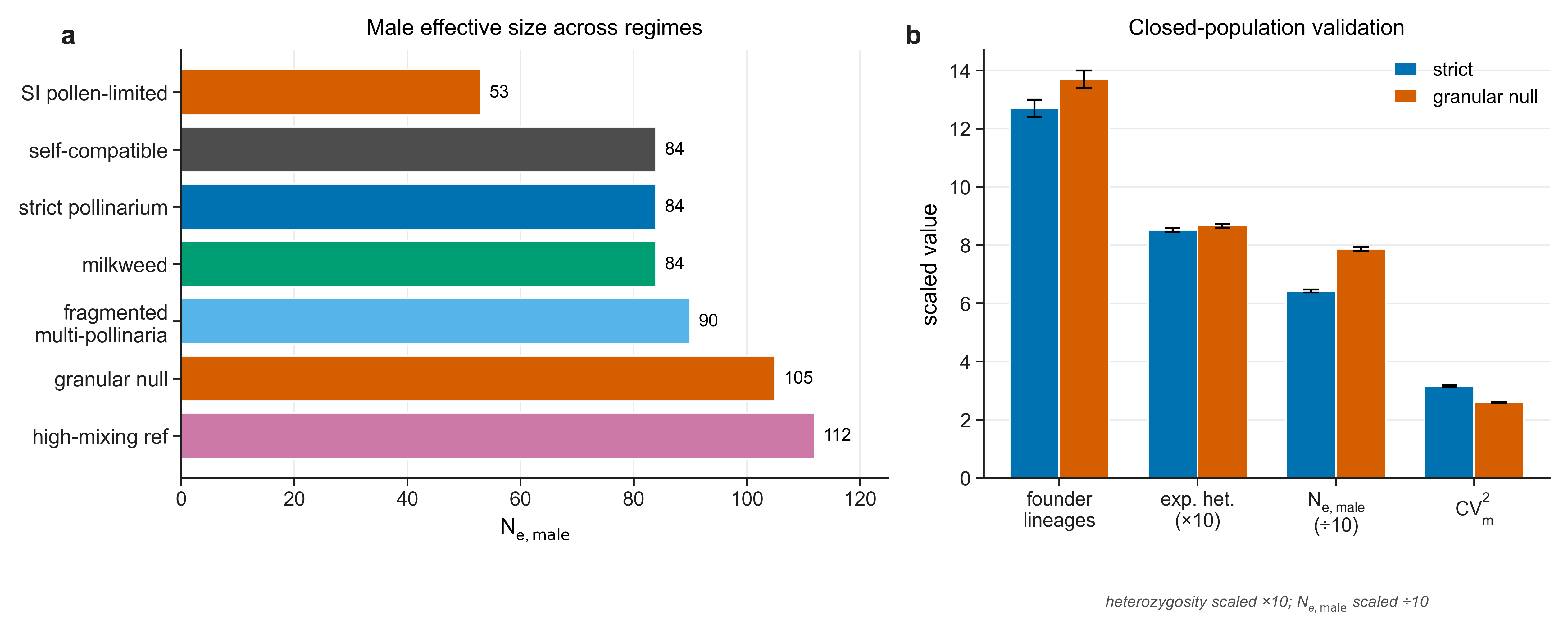
Seven, from strict pollinarium and fragmented multi-pollinaria through milkweed paired transfer with late-acting self-incompatibility to a high-mixing reference.
Parameter ranges are literature-anchored, with deposition sets for named taxa tuned to observed samples and screened by global sensitivity analysis.
A variance effective size computed from the realised offspring-count distribution for each sex channel, plus founder-lineage retention.
A two-layer result
Within the parameter envelope explored, packaged transfer leaves a clean two-layer signature.
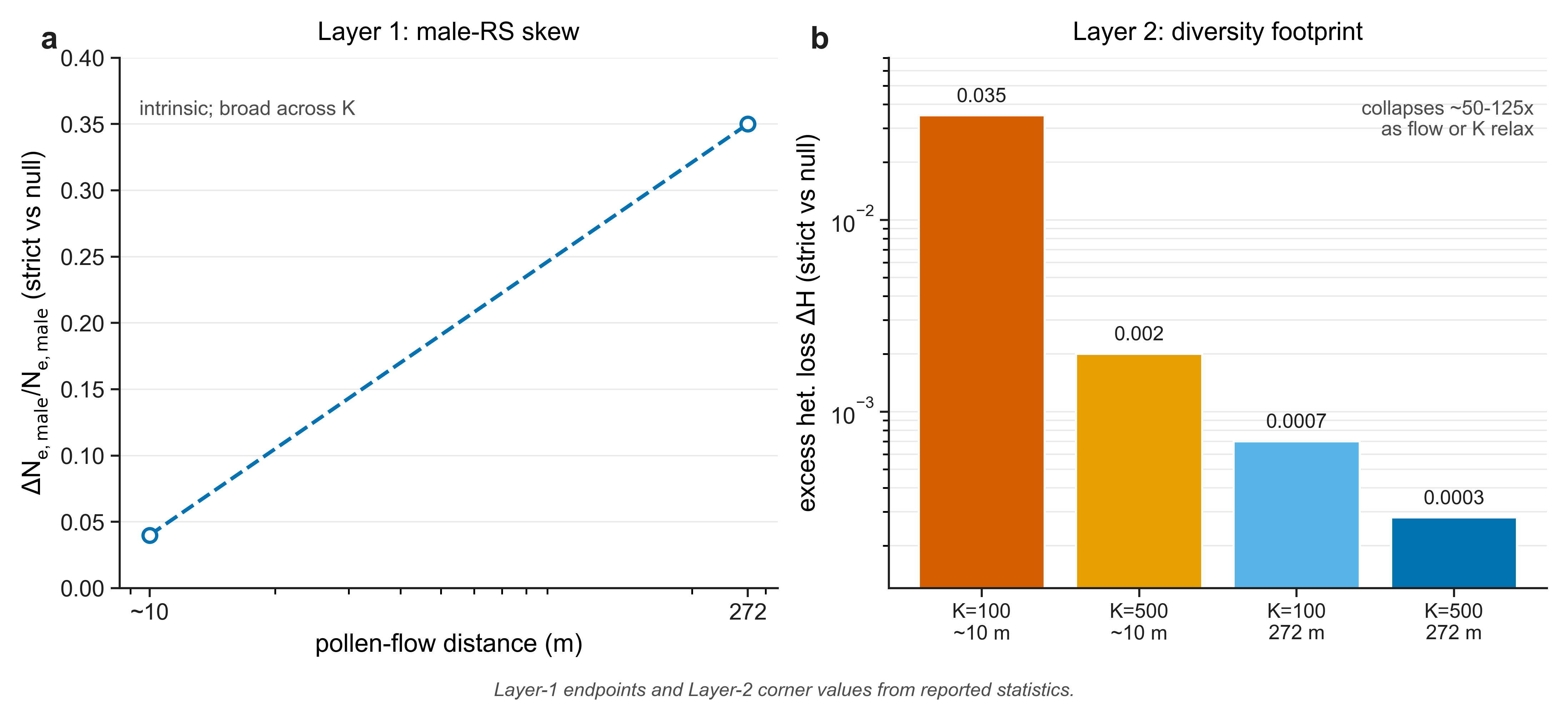
- Male reproductive-success variance is intrinsic and broad. In the single-sire regime, packaging depresses male effective size relative to the granular null by roughly a tenth to a third — and, counter-intuitively, the contrast is clearest under broad pollen flow.
- The diversity footprint is demographically gated. Any population-genetic effect on diversity is confined to small, fragmented, short-pollen-flow populations, and collapses by one to two orders of magnitude once gene flow or census size relax.
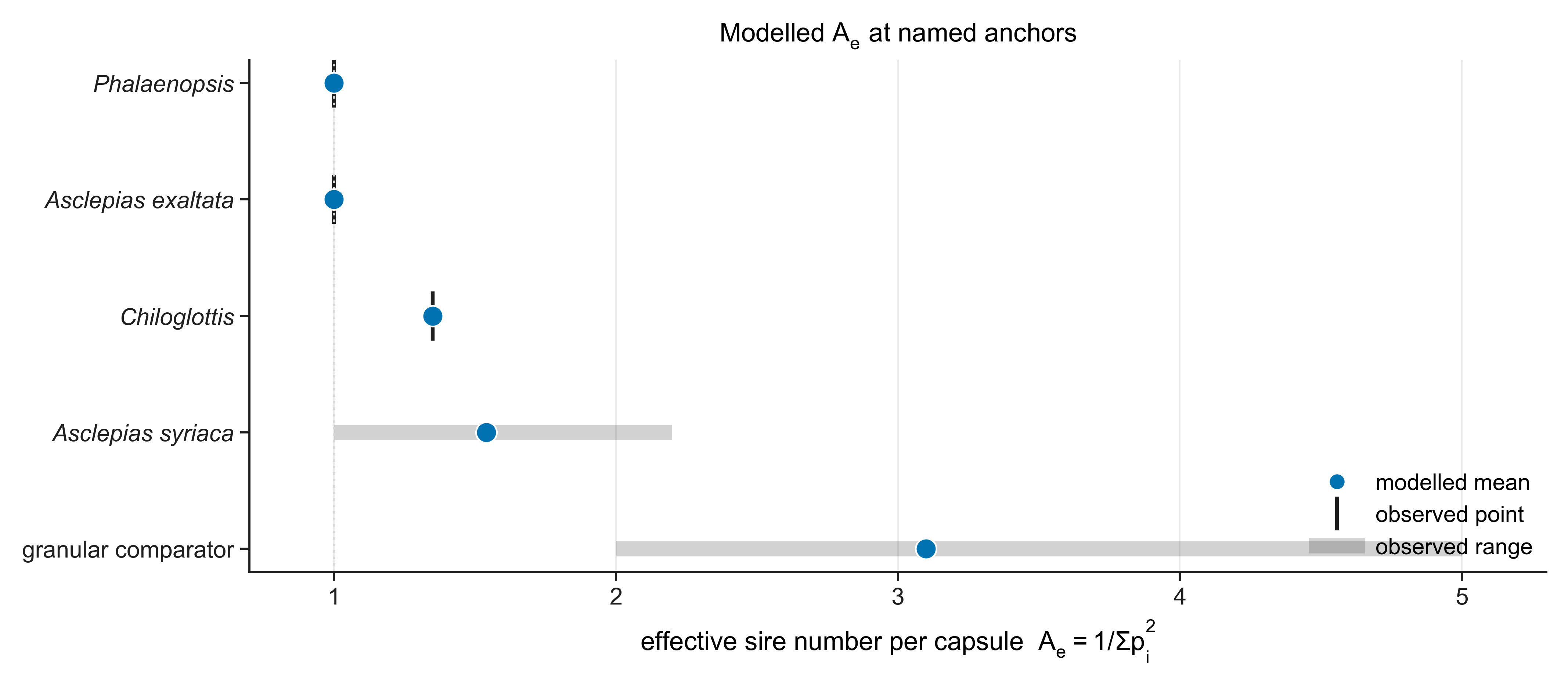
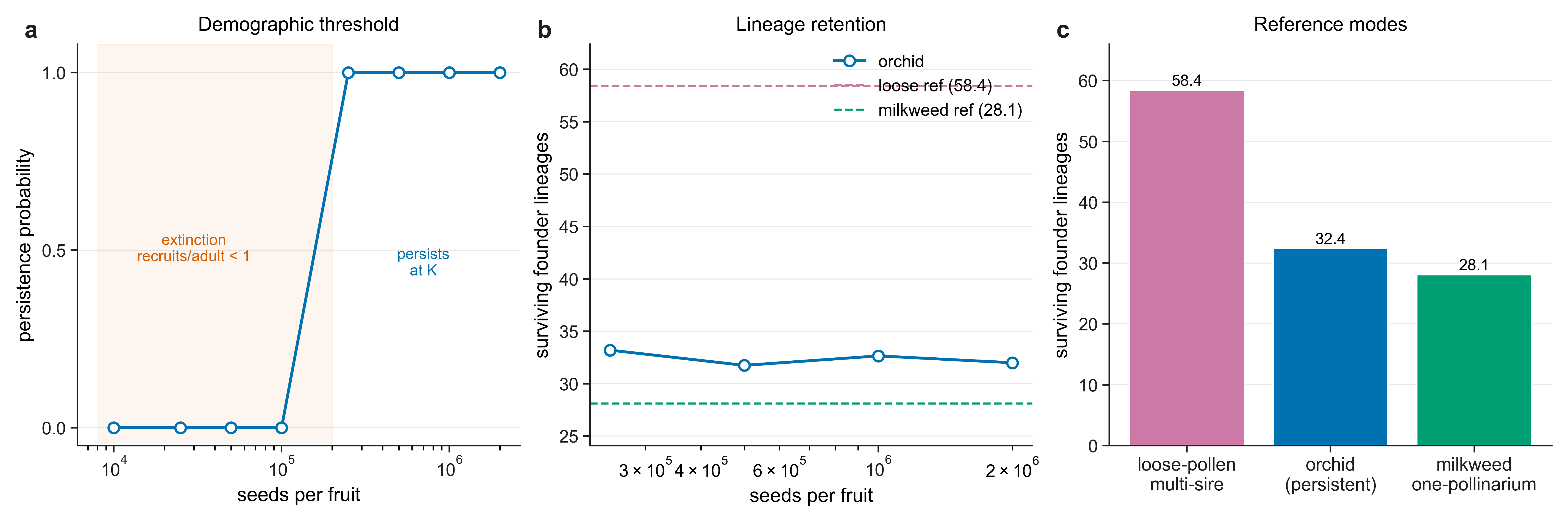
Placed against reference systems, an orchid that sweeps a whole capsule with one pollinarium is as single-sire as Asclepias, yet — through sheer seed and fruit output — lifts founder-lineage retention off the milkweed floor toward, but not up to, a loose-pollen multi-sire reference.
Where the consequence really lives
Pollinaria do create a distinctive paternity structure with measurable genetic consequences. But those consequences live in the second moment of the male fitness distribution — the variance of male reproductive success — and inside a namable demographic envelope, not in a general erosion of population diversity. The deterministic "packaging destroys diversity" story does not survive a properly matched null.
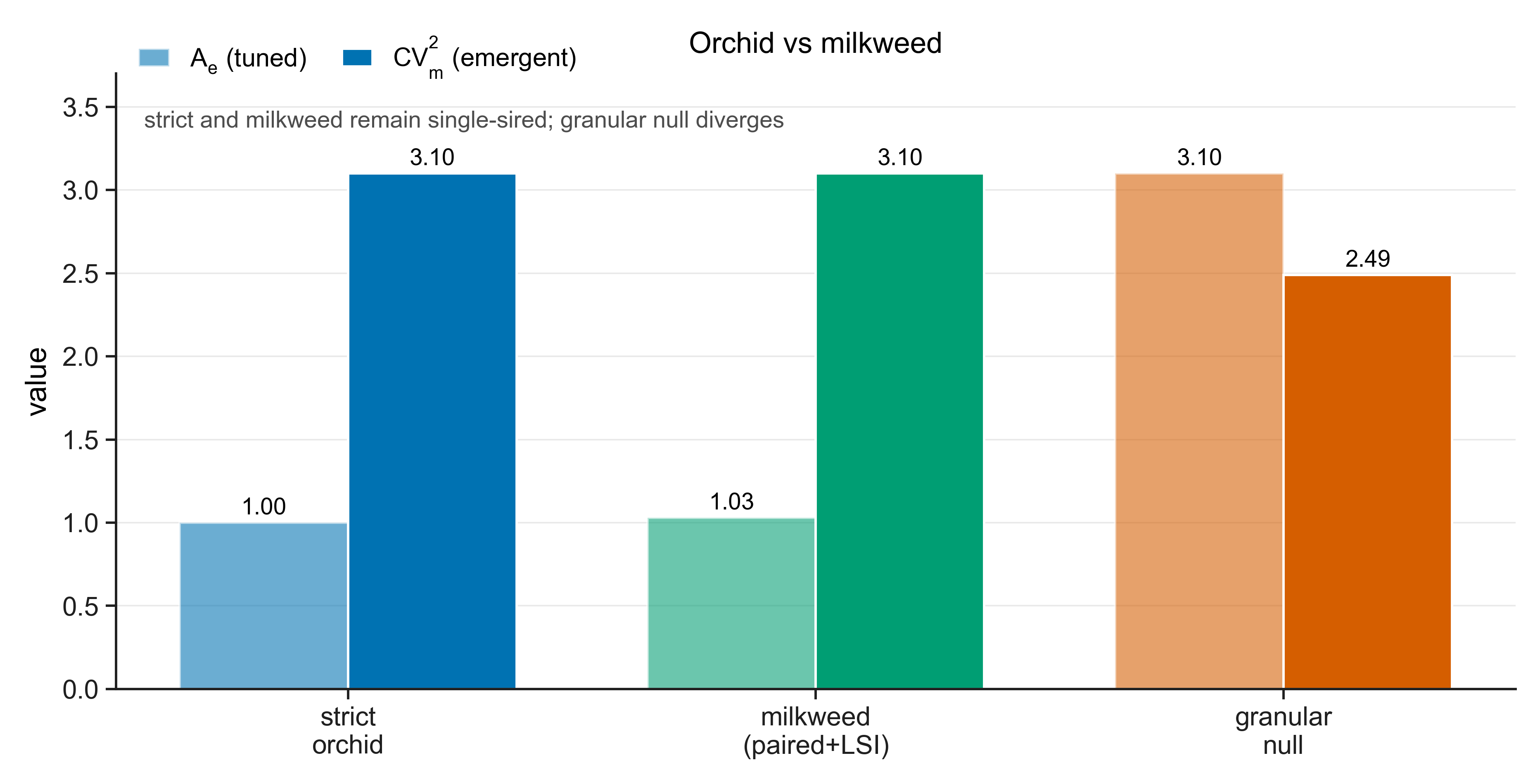
A speciation hypothesis
Stated explicitly as hypothesis rather than result: a single-sire system could reward a rare male-function innovator with whole capsules rather than a diluted share. That would supply an economic precondition for budding speciation along the placement axis opened by the viscidium — a female-derived adhesive that shares a developmental backbone with the stigma, and the subject of Orchidarc's parallel biomaterials programme.
Evidential discipline. The manuscript carries an explicit source-verification table separating background, empirical anchors, model parameters and speculation, so that no claim silently migrates from background to evidence.
Manuscript
Single-sire capsules in polygamous flowering plants: pollinaria as the zero-carryover endpoint of the pollen-transfer continuum and a conditional driver of male reproductive skew. Andrés E. Ramos R. Manuscript in preparation.
Research themes
- The pollen-carryover continuum and the pollinarium as its zero-carryover endpoint.
- Capsule-level single paternity and effective sire number as emergent deposition properties.
- Male reproductive-success variance under packaged versus granular pollen transfer.
- Demographic gating of diversity effects in fragmented orchid and milkweed populations.
- Placement, the viscidium and the economics of male-function innovation.