
The orchid pollinarium is often described as an anatomical innovation. It is also a mechanical device. It must attach in seconds, survive transport on a moving animal, then release pollen only when a second flower's stigma provides a competing grip.
The literature describes who pollinates orchids and when, but rarely how the pollinia are physically transferred or why only some visits succeed. This programme asks that mechanical question directly. Its first study takes Cattleya walkeriana (Laeliinae) and measures what happens during a single extraction event — connecting Orchidarc's two material systems, the viscidium that attaches the pollinarium to a vector and the stigmatic composite that captures and retains the pollen mass.
The pollination sequence
Orchid pollination can be treated as a sequence of coupled mechanical events. Each event has a different dominant material requirement, and failure at any stage can prevent fertilisation.
The pollinator touches the viscidium or an associated adhesive surface during floral entry or manipulation.
The viscidium wets, bonds or mechanically locks onto the pollinator surface.
The pollinarium must remain attached while the animal moves, vibrates, grooms or contacts other surfaces.
The pollinia enter the stigma and are retained by the stigmatic matrix during withdrawal.
A flower as a mechanical system
In Cattleya walkeriana the pollinarium is typical of Laeliinae: four pollinia linked directly to one another by appendicular caudicles, a viscidium at the base, and no stipe. The stigma is not a convex surface but a deep concavity — here roughly 6.5 mm across and 3 mm deep — filled with the cell-rich stigmatic composite into which the pollinator presses the pollinia as it withdraws.
Successful deposition therefore hinges on two coupled events: the stigmatic composite must hold the pollinia against the tension of the retreating animal, and the caudicle–pollinium interface — the designated weak point of the appendicular architecture — must fracture before the whole pollinarium is pulled free. Whether that release happens cleanly depends on the rate at which the load is applied, because both the caudicle and the gel are viscoelastic. The connecting matrix itself, elastoviscin, is a lipid-rich tapetal material that can stretch a caudicle to more than ten times its resting length before breaking.
Reproducing the extraction event
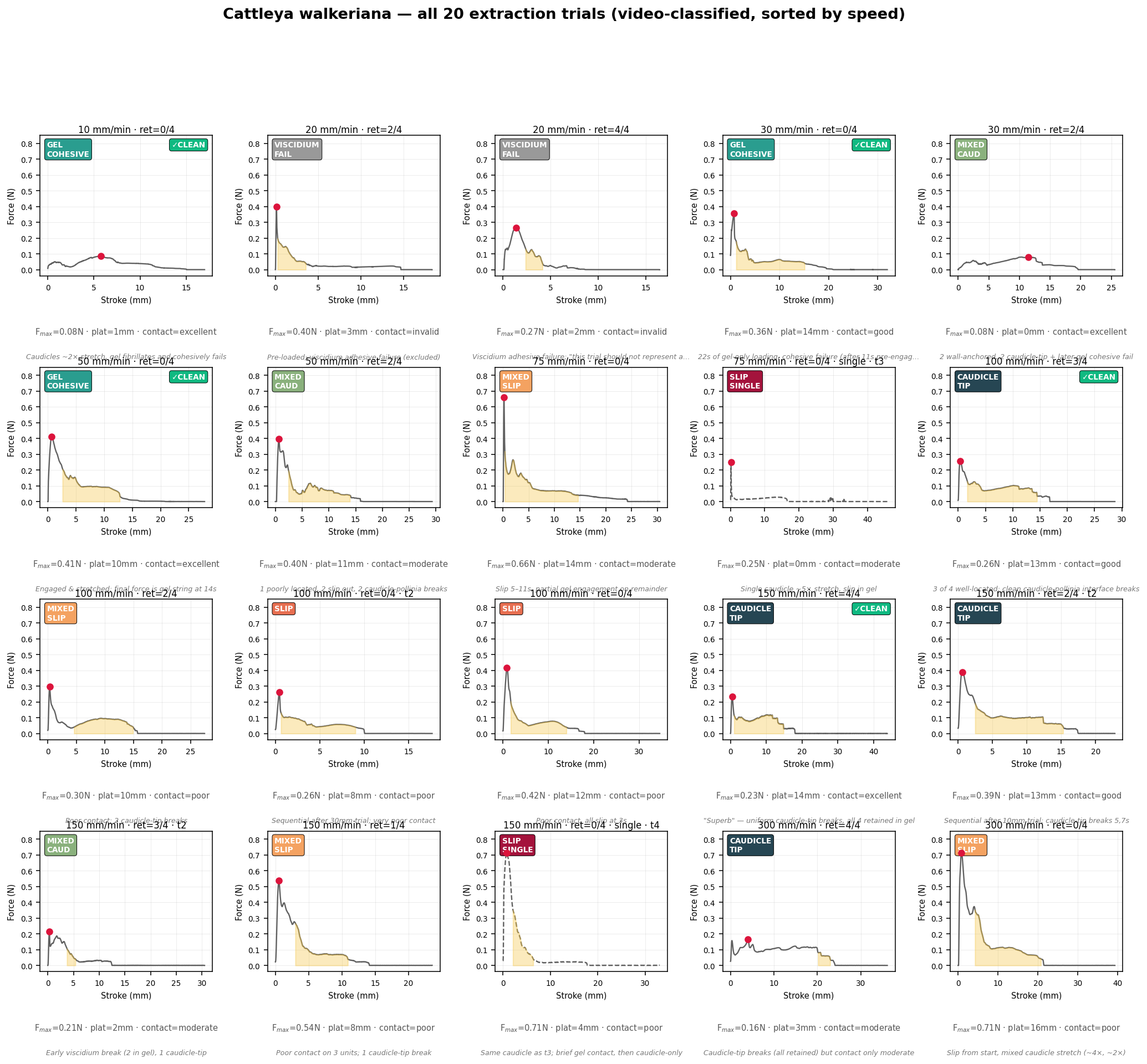
A 3D-printed pollinator mimic mounted on a tensile-testing apparatus reproduces the withdrawal under controlled conditions. A 10 N load cell records force and crosshead displacement at 100 Hz, synchronised to a 30 fps lateral video so that every mechanical event can be matched frame-by-frame to the force trace. Twenty extraction trials spanned a thirtyfold range of withdrawal speeds (10–300 mm min⁻¹).
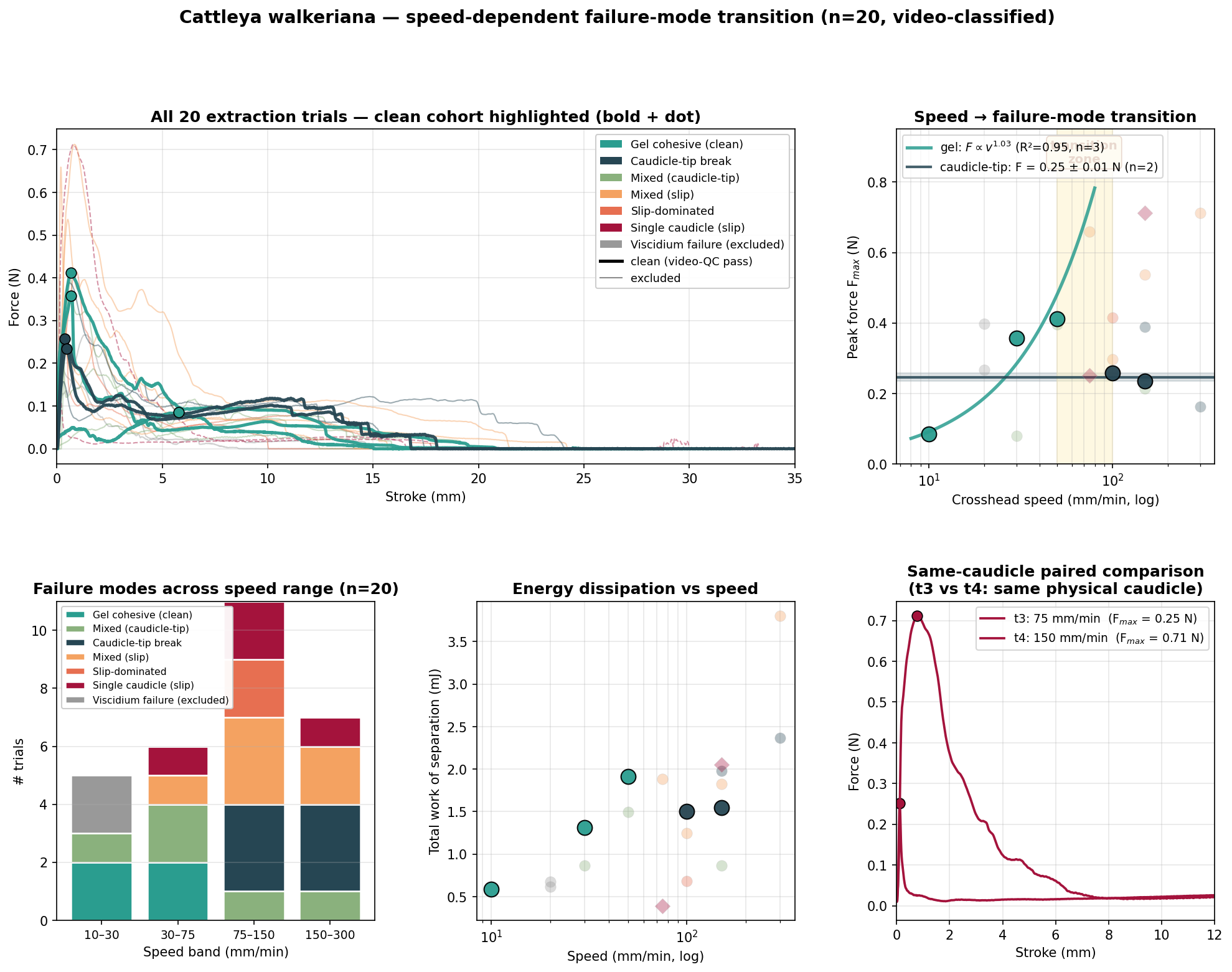
Crucially, each trial was classified by its video-observed failure mode, independent of the recorded force. A stringent contact-quality criterion — good gel contact, all four pollinia engaged, a single dominant failure mode — left a five-trial "clean" cohort that carries the headline result.
Peak force, work of separation and multi-peak events at 100 Hz, baseline-corrected for column self-weight.
30 fps record used to classify gel fibrillation, caudicle rupture, slip and viscidium detachment.
Optically measured cross-section and gauge length, converting stiffness to a tangent modulus.
Two regimes, one threshold
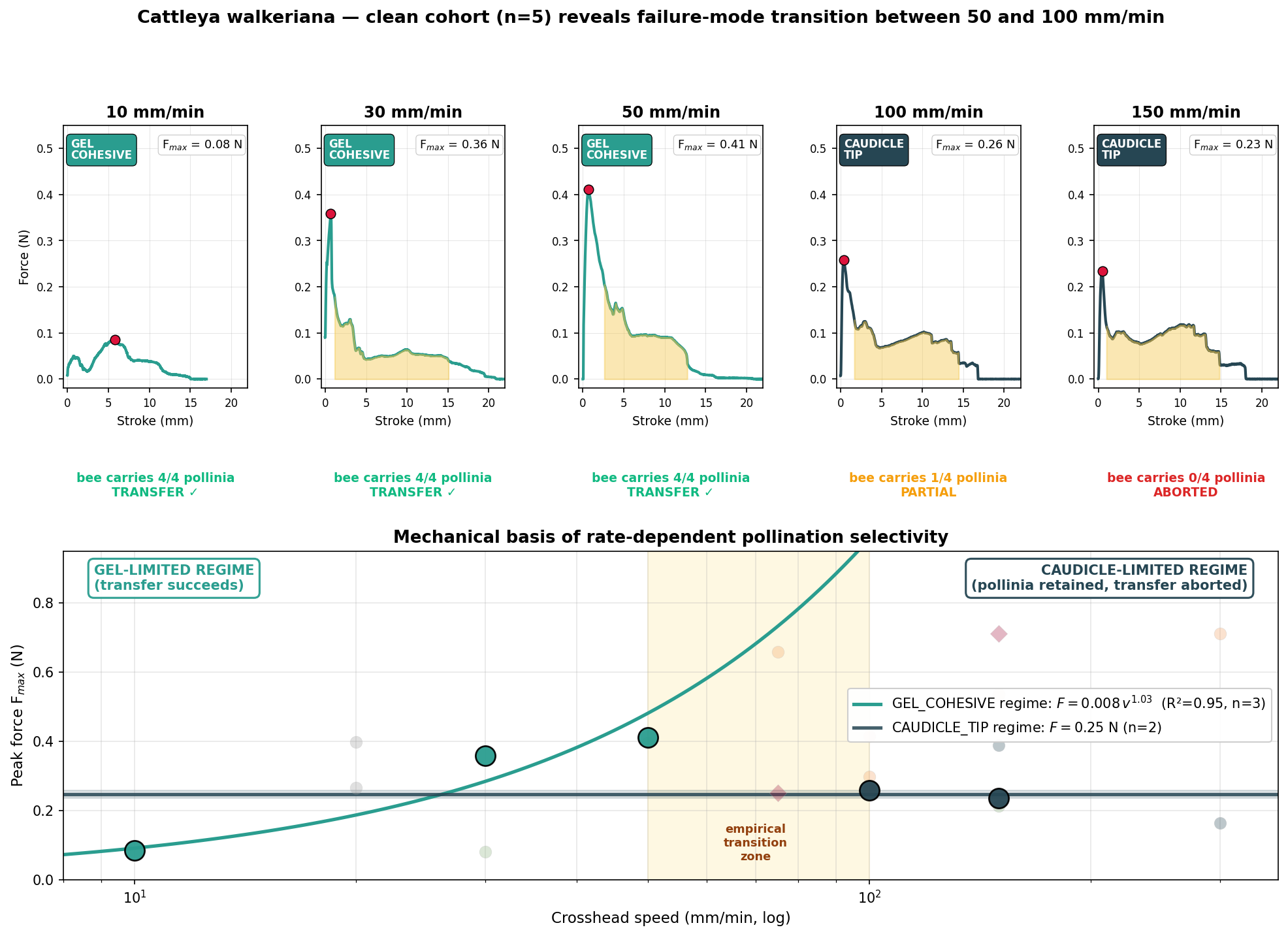
The clean cohort shows a categorical change in mechanism between 50 and 100 mm min⁻¹. The same column, the same gel and the same pollinia produce opposite outcomes either side of that boundary.
At 10–50 mm min⁻¹ the stigmatic composite fibrillates and ruptures internally; caudicles stretch only modestly and the pollinia leave fully embedded in the mimic. Complete pollen transfer.
At 100–150 mm min⁻¹ the gel holds and the caudicle–pollinium interface fails instead; the mimic departs with bare caudicles and no pollen. Failed transfer.
Within the slow regime, peak force scales almost linearly with speed (Fₘₐₓ = 0.0085·v¹·⁰³ N, R² = 0.95) — the signature of a viscoelastic gel. The caudicle-tip threshold, by contrast, is rate-independent at ~0.25 N, the mark of a brittle interfacial bond. Which interface fails — and therefore whether the bee leaves with the pollinia or without them — is set by the competition between these two laws.
The core result. The pollinarium is not a passive pollen trap. It is a tunable two-parameter mechanical filter: a rate-dependent gel-cohesive force versus a rate-independent caudicle-tip threshold, whose relative magnitudes define a species-specific window of withdrawal rates compatible with pollen export.
Caudicles across genera
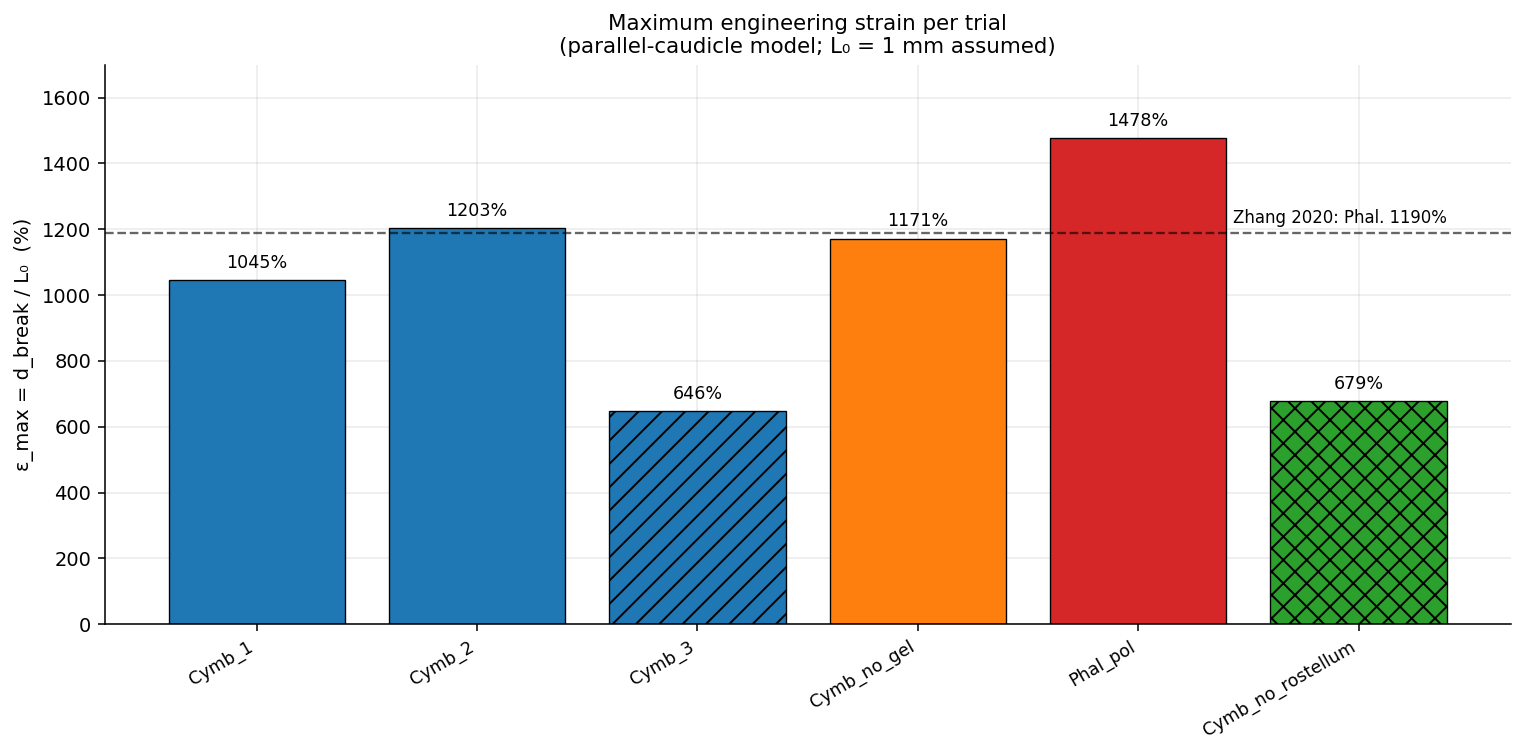
The same apparatus, applied to other genera, places C. walkeriana on a cross-genus stiffness spectrum. Its in-cavity caudicle tangent modulus is 33–44 kPa — the compliant end — against Phalaenopsis at ~152 kPa and Cymbidium at ~286 kPa. The Phalaenopsis value agrees to within 5% with an independent single-fibre benchmark (Zhang et al. 2020, ~160 kPa) measured by a completely different method, cross-validating the in-cavity approach.
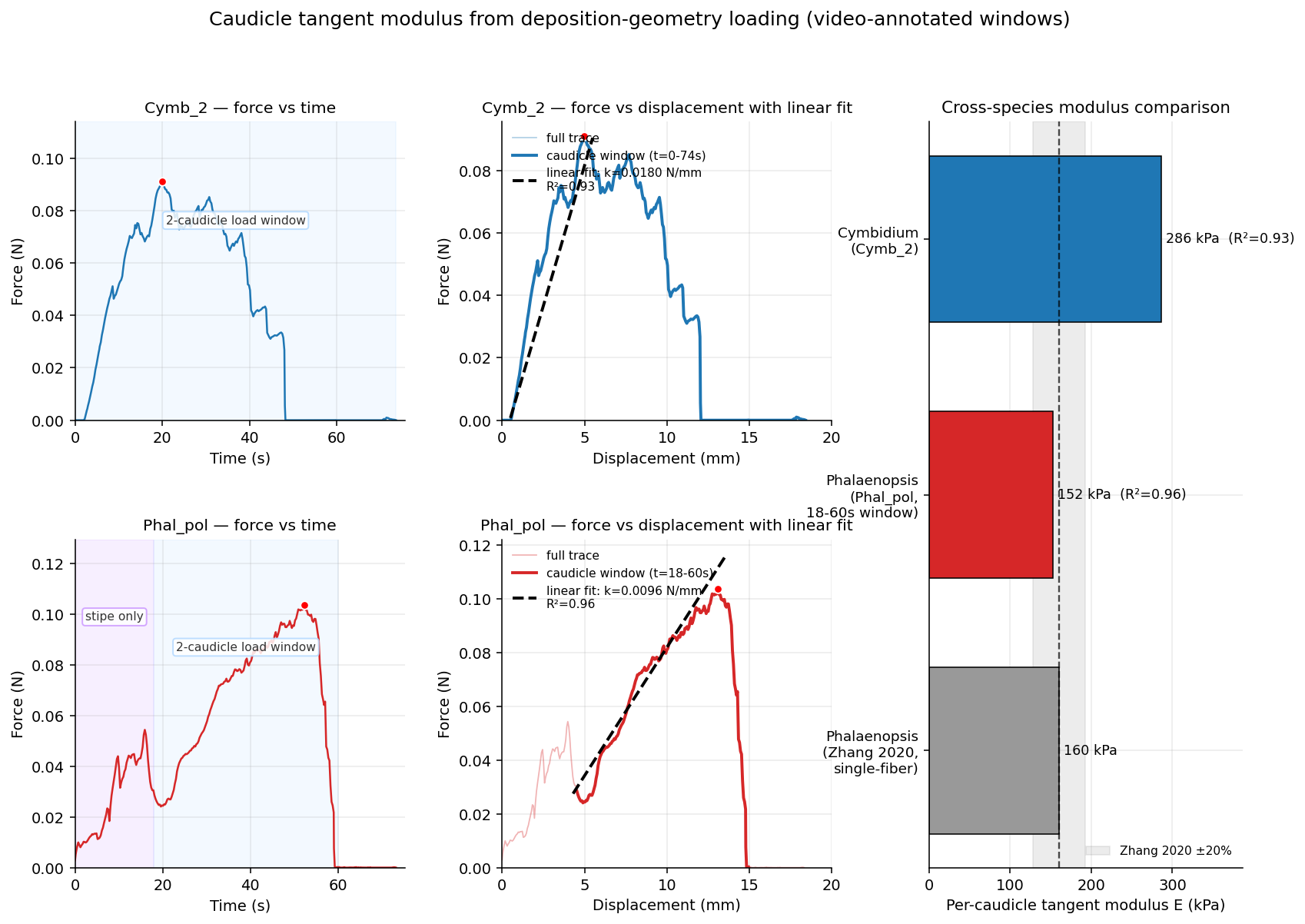
Despite that fourfold difference in stiffness, the C. walkeriana breaking strains (836% and 1196%) fall squarely in the range reported for vandoid caudicles — the first quantitative caudicle-strain values for the appendicular-caudicle clade, and evidence that extreme extensibility is conserved across the appendicular–frenicular architectural divide.
A mechanical filter for pollinator behaviour
A system that flips outcome across less than a fourfold change in rate is, by construction, a filter for pollinator behaviour: decisive departures and prolonged contact produce categorically different results. C. walkeriana is pollinated by male euglossine bees, and whether the filter selects for or against transfer at their natural withdrawal rates is set by the relative magnitudes of the gel-cohesive force and the caudicle-tip strength — both quantitative material parameters that selection can tune.
This gives a physical basis for the broader hypothesis that orchid pollinator specificity is partly enforced by stigmatic-composite rheology, complementing the chemical specificity of floral fragrance and the morphological specificity of column and labellum. It also connects directly to the frequency-dependent stiffening measured in the stigmatic composite programme, where Cattleya was the one genus whose storage modulus stayed above the Dahlquist tack threshold across the whole frequency window.
Limitations and next steps
The headline rests on a small clean cohort: three trials anchor the gel-cohesive force law and two anchor the caudicle-tip threshold. The transition is categorical in every contact-clean trial, but contact quality — judged from video — was the dominant source of trial-to-trial variability, and only five of twenty trials passed the criterion.
A positioning jig and randomised trial order to raise the yield of contact-clean trials per speed.
Three or more trials at each of 30, 50, 100 and 150 mm min⁻¹ to firm up both mechanical laws.
Carry the modulus and failure-mode analysis to Cymbidium, Dendrobium and other pollinarium-bearing taxa.
Manuscript
Rate-dependent failure-mode selection in the pollinarium of Cattleya walkeriana: a mechanical filter for pollinator behaviour. Andrés Ramos [co-authors to confirm]. Preliminary draft.